Álvaro García
Few nutritional concepts in dairy production generate as much discussion, and as much misunderstanding, as fiber. Within formulation software, fiber is typically presented as a quantitative fraction, expressed through percentages, digestibility coefficients, and predicted production responses. This representation is convenient, mathematically tractable, and operationally necessary. Yet it also encourages a reductionist view in which fiber is treated primarily as a chemical entity rather than a functional regulator of rumen physiology.
In the rumen, fiber does not behave as a passive nutrient pool. It operates as a dynamic regulatory force acting through both mechanical and metabolic pathways. Structural characteristics influence chewing behavior, saliva secretion, and digesta kinetics, while fermentability governs microbial activity, volatile fatty acid production, and acid load dynamics. These processes are inseparably linked yet mechanistically distinct. When fiber is evaluated solely through concentration metrics, the biological complexity governing rumen stability is easily obscured.
Much of the persistent confusion surrounding fiber nutrition arises from the collapse of these fundamentally different mechanisms into a single nutritional narrative. Chemical adequacy is frequently interpreted as functional adequacy, and fermentability is often conflated with physical effectiveness. As a result, diets that appear balanced on paper may generate responses that seem inconsistent or contradictory under field conditions. The challenge, therefore, is not simply quantifying fiber, but understanding how fiber functions within the rumen system.
Neutral detergent fiber describes a chemical fraction. Physically effective fiber describes a functional property. Fermentable fiber describes a metabolic substrate. These terms are often used interchangeably in conversation, yet they represent different dimensions of rumen physiology. Failure to recognize these distinctions remains a recurring source of nutritional instability and misinterpreted responses.
Physically effective fiber exerts its primary influence through mechanical stimulation of the rumen. Particle size, structural integrity, and resistance to rapid degradation determine the extent to which a feed stimulates chewing and rumination. These processes govern saliva secretion, and saliva remains the cow’s dominant buffering system. Through its bicarbonate content, saliva moderates’ fermentation acid load, stabilizes rumen pH, and sustains the motility patterns required for efficient digesta turnover. This is not a secondary effect layered onto fermentation; it is a central control mechanism.
Highly fermentable fiber sources such as soyhulls and beet pulp operate through an entirely different route. Their contribution arises primarily from fermentation dynamics. These feeds provide substrates that modify microbial activity, alter volatile fatty acid production, and shift VFA molar proportions. Such shifts can influence rumen pH, yet the mechanism is metabolic rather than mechanical. Fermentation-driven regulation does not replicate chewing-driven buffering.
A diet rich in fermentable fiber may enhance energy supply while providing limited stimulation of rumination. From a chemical standpoint, fiber concentration may appear entirely adequate, satisfying formulated NDF targets and predicted digestibility parameters. Yet rumen physiology is regulated not only by nutrient composition but also by physical stimuli. When fiber is supplied primarily through finely processed ingredients or highly digestible byproducts, the structural signals required to drive chewing activity may be weakened.
Both pathways influence rumen function, but they are not interchangeable
This distinction is critical. Rumination is not simply a behavioral response but a vital component of rumen homeostasis. Chewing activity governs saliva secretion, and saliva represents the cow’s dominant buffering mechanism against fermentation-derived acid load. Reduced rumination alters buffering dynamics, digesta stratification, and rumen motility patterns. These changes may occur even when fermentable substrates are abundant and predicted energy supply remains high.
The consequence is a physiological disconnect between chemical adequacy and functional effectiveness. Rations may appear balanced analytically yet fail to generate the mechanical stimulation necessary to stabilize rumen conditions. This helps explain why diets formulated with high inclusion rates of finely ground feeds, pelleted ingredients, or rapidly fermentable fiber sources sometimes produce responses consistent with structural fiber deficiency. Variability in rumen pH, altered intake behavior, milk fat depression, and increased sensitivity to management disturbances frequently emerge under these conditions despite acceptable NDF values.
In practical terms, fiber concentration alone does not guarantee rumen stability. Functional fiber effectiveness remains the decisive factor.
Rumen stability is governed by interacting forces. Acid production reflects fermentation intensity and carbohydrate availability. Buffering capacity reflects salivary flow and rumination dynamics. Digesta kinetics reflect both fermentation rate and physical structure. Intake regulation reflects the interaction of metabolic feedback and rumen fill constraints. No single variable dictates system behavior. Stability emerges from the balance among mechanisms operating simultaneously. Reducing this system to fiber percentage oversimplifies biological reality.
Milk fat responses provide a clear example of this interaction. Physically effective fiber supports rumination, saliva production, and rumen conditions favoring acetate-dominant fermentation. Acetate production is strongly associated with milk fat synthesis. Fermentable fiber contributes fermentable substrate, yet without sufficient mechanical stimulation, the rumen environment itself may shift toward patterns that alter fermentation efficiency and acid load dynamics.
The response is therefore governed not solely by substrate supply, but by regulatory balance. Equating fermentable fiber with physically effective fiber risks overestimating the stabilizing capacity of highly digestible feeds. Fermentable fiber contributes energy and modulates fermentation patterns. Physically effective fiber governs rumination intensity and buffering mechanics. These functions intersect but do not substitute for one another.
When imbalance occurs, the biological consequences may manifest as milk fat depression, altered intake behavior, rumen pH variability, or increased sensitivity to management disturbances. Such outcomes are often attributed to ingredients rather than underlying mechanisms.
Effective ration formulation therefore requires a multidimensional perspective. Fiber must be evaluated not only by concentration but by physical effectiveness, particle distribution, degradation characteristics, and interaction with fermentation kinetics. Diets optimized chemically but misaligned physically may exhibit instability even when predicted values suggest adequacy.
Physically effective fiber and fermentable fiber should be understood as complementary regulators within the rumen ecosystem. One governs mechanical buffering and motility dynamics. The other governs fermentation patterns and substrate availability. System efficiency depends on their interaction rather than their substitution.
Fiber is not merely a nutrient fraction. It is a structural, mechanical, and metabolic driver of rumen regulation. Precision feeding strategies must therefore recognize that fiber functionality, not fiber concentration alone, determines stability, efficiency, and production consistency. The distinction is not theoretical. It is operational.
Rethinking peNDF: A mechanistic inflection between fermentative efficiency and physical constraint
The response of high-producing dairy cows to increasing peNDF cannot be adequately explained by changes in rumination time or shifts in VFA molar proportions alone. Rather, peNDF operates within an integrated regulatory system in which mechanoreceptor-driven physical signals and chemoreceptor-mediated metabolic feedback jointly determine intake, fermentation kinetics, and productive efficiency. The productive outcome of fiber inclusion is therefore nonlinear and characterized by a biological inflection point. At low peNDF supply, ruminal regulation is dominated by chemical instability. Reduced particle length and insufficient fiber rigidity diminish mechanoreceptor stimulation, lowering rumination and saliva secretion. As buffering capacity declines, rapid fermentation of non-fiber carbohydrates elevates total VFA concentration and shifts molar proportions toward propionate. Rumen pH decreases, microbial ecology becomes destabilized, and chemoreceptor signaling intensifies. Under these circumstances, incremental increases in peNDF restore rumination-mediated buffering, temper acid accumulation, and reestablish microbial conditions conducive to sustained fermentative activity. Importantly, the benefit of peNDF at this stage is not merely an increase in acetate proportion, but the preservation of total VFA production under physiologically stable conditions. Here, additional peNDF enhances both rumen integrity and productive output.
Beyond a critical threshold, however, regulatory dominance shifts. As indigestible or slowly fermentable fiber accumulates, rumen distension increases relative to metabolizable substrate supply. Mechanoreceptor signaling begins to constrain intake independent of metabolic demand. In this state, ruminal regulation transitions from chemically mediated to physically constrained. Although acetate proportion may continue to rise and pH may appear favorable, total fermentable organic matter declines, fermentation rate slows, and absolute VFA production is reduced. The system exhibits apparent chemical stability while simultaneously losing energetic efficiency. This inflection is particularly consequential in early lactation.
High-producing cows are metabolically prioritized for glucose supply and rely heavily on propionate-driven gluconeogenesis. When excessive peNDF displaces fermentable carbohydrate, the decline in total VFA yield and microbial protein synthesis reduces metabolizable energy availability at a time when intake is already limiting. The result is a paradoxical scenario: milk fat concentration may be preserved or increased, yet energy-corrected milk declines because fermentative throughput has been physically restricted. The critical insight is that optimal peNDF is not defined by maximal rumination, maximal acetate proportion, or even maximal rumen pH. It is defined by the point at which fermentative throughput, expressed as total VFA production and microbial biomass yield, is maximized without triggering acidotic feedback or intake suppression. Below this point, chemical risk predominates and peNDF provides corrective value. Above it, physical limitation predominates and additional structural fiber imposes an energetic penalty.
Thus, highly indigestible fiber sources such as wheat straw should be conceptualized as regulatory modifiers rather than production drivers. Their role is to prevent destabilization of the ruminal environment, not to anchor the structural foundation of high-producing diets. Precision feeding in modern dairy systems requires recognition that rumen stability and fermentative efficiency are not synonymous, and that maximizing one at the expense of the other defines the biological cost of exceeding the peNDF inflection point.
Figure 1 below illustrates a conceptual model describing the biphasic response of high-producing dairy cows to increasing peNDF. This model illustrates the nonlinear relationship between dietary peNDF inclusion and fermentative efficiency in high-producing dairy cows. The x-axis represents increasing peNDF concentration (with distinction between digestible and indigestible sources), and the y-axis represents productive efficiency, defined as total volatile fatty acid (VFA) production and energy-corrected milk yield under stable rumen conditions. In Phase I (chemical regulation domain), low peNDF supply results in reduced rumination, diminished saliva secretion, and compromised ruminal buffering. Rapid fermentation elevates total VFA concentration, shifts molar proportions toward propionate, and decreases rumen pH. Chemoreceptor-mediated feedback predominates, increasing acid load risk and destabilizing microbial ecology. Incremental increases in peNDF within this scenario enhances mechanoreceptor stimulation, restore rumination, stabilize rumen pH, and support sustained total VFA production. Productive efficiency increases as acidogenic risk is mitigated. At the inflection point, peNDF supply achieves sufficient mechanoreceptor stimulation to maintain rumen stability without imposing physical intake constraints.
Total VFA production and microbial protein synthesis are maximized under stable pH conditions. This point represents the optimal balance between fermentative throughput and structural adequacy. In Phase II (physical regulation domain), further increases in peNDF, particularly from low-digestibility sources, shift regulatory dominance to mechanoreceptor-mediated rumen fill limitation. Intake becomes constrained by physical distension rather than metabolic demand. Although ruminal pH remains stable and acetate molar proportion may increase, total fermentable organic matter declines, fermentation rate decreases, and absolute VFA production is reduced. Productive efficiency declines due to energy dilution and reduced metabolizable substrate supply. The model emphasizes that optimal peNDF concentration is not defined by maximal rumination time or maximal acetate proportion, but by maximal fermentative throughput achieved without triggering acidotic feedback or intake suppression. Exceeding this threshold results in a biologically predictable trade-off between rumen stability and energetic efficiency.
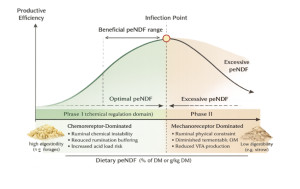
Figure 1. Biphasic response of productive efficiency to increasing dietary physically effective NDF (peNDF).
Conclusions
Fiber evaluation in dairy nutrition must move beyond concentration metrics toward functional interpretation. Neutral detergent fiber, physically effective fiber, and fermentable fiber represent distinct yet interacting regulators of rumen physiology. Treating these dimensions as interchangeable obscures the biological mechanisms governing rumen stability, intake regulation, and productive efficiency.
Physically effective fiber operates primarily through mechanical pathways, modulating rumination, salivary buffering, digesta kinetics, and ruminal stratification. Fermentable fiber functions through metabolic pathways, shaping microbial activity, VFA production, and energy supply. Although both influence rumen conditions, their mechanisms are not substitutable. Chemical adequacy does not guarantee functional effectiveness.
The response to increasing peNDF is inherently nonlinear. At low inclusion levels, insufficient physical stimulation compromises buffering capacity and microbial stability, constraining fermentative efficiency. Incremental increases restore rumen homeostasis and maximize fermentative throughput. Beyond a biological inflection point, however, excessive peNDF, particularly from low-digestibility sources, imposes rumen fill limitations that reduce fermentable substrate availability, total VFA yield, and productive efficiency despite apparent pH stability.
Optimal fiber supply is therefore defined not by maximal rumination, maximal acetate proportion, or maximal rumen pH, but by maximal fermentative throughput achieved without triggering acidotic feedback or physical intake constraints. Rumen stability and energetic efficiency are complementary but not synonymous objectives.
Precision feeding strategies must recognize fiber as a structural, mechanical, and metabolic regulator rather than a passive nutrient fraction. Incorporating functional fiber dynamics into ration formulation is essential for improving production consistency, mitigating nutritional instability, and aligning diet design with rumen biology.
The full list of references used in this article is available upon request.
© 2026 Dellait Knowledge Center. All Rights Reserved.





